|
Verbascum thapsus
|
|
|
|
Scientific name
|
Verbascum thapsus
|
|
Additional name information:
|
L.
|
|
Common name
|
common mullein, wooly mullein, great mullein, mullein, JacobÛªs staff, flannel leaf, velvet plant, candlewick plant, lung wort, felt wort
|
|
Synonymous scientific names
|
none known
|
|
Closely related California natives
|
0
|
|
Closely related California non-natives:
|
3
|
|
Listed
|
CalEPPC List B,CDFA nl
|
|
By:
|
Michael Pitcairn
|
|
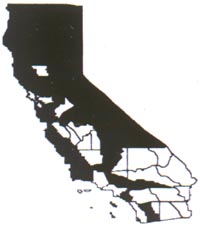
Distribution
|
|
HOW DO I RECOGNIZE IT?
Distinctive features:
|
Common
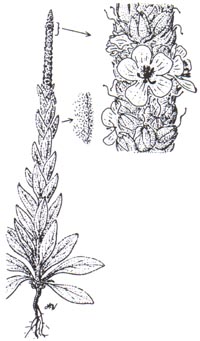
mullein (Verbascum thapsus ) is a biennial or annual herb that sends up a large stalk, three to six feet tall, topped with yellow flowers closely attached to the stalk. It spends the first year as a rosette close to the ground. The leaves are large, six to twelve inches, densely woolly, and soft to the touch. Leaves are largest at the base and gradually become smaller up the stalk. Leaves on the stalk occur alternately, and the base of each leaf extends a short way down the stalk. Yellow flowers, three-quarters to one and a half inches in diameter, consist of five circular petals, and occur on the uppermost portion of the stalk. |
|
|
|
Description:
|
Scrophulariaceae. Biennial or annual herb.
Stems: 1.5-6 ft (50-200 cm) tall when mature. Leaves: first-year plant is a
rosette of large, woolly, gray-green leaves. Leaves 2-4 times longer than wide,
2-16 in (5-40 cm) long, alternate, the largest leaves at the base of the plant,
smallest near the top; leaf pedicle short with leaf base extending a short way
down the stalk to form wings. Plants usually bolt in second year and have a
single stem covered with overlapping, woolly leaves from base to inflorescence.
Inflorescence: a spike extending upward from top of stalk. Usually 1
inflorescence is produced, but occasionally a second or third can form with
branches occurring where inflorescences begin. Flowers: densely packed along
inflorescence, youngest near top; bracts 0.5-0.7 in (12-18 mm), flower pedicels
short,
åÊ
|
|
WHERE WOULD I FIND IT?
|
Common mullein occurs throughout California,
but is particularly abundant in dry valleys on the eastern side of the Sierra
Nevada. High population densities have been observed in moist meadows and creek
drainages near Mono Lake and Owens Valley. It prefers disturbed habitats with
little other vegetation, especially on dry, gravelly soils. It is common along
roadsides, rights-of-way, and river banks and in forest cuts, meadows, pastures,
and waste areas (Gross and Werner 1978). It is an early colonizer and may be the
first plant to colonize bare soil. It is found in all forty-eight contiguous
states and in Hawaii. In Canada it is reported to grow abundantly in soils with
a pH range of 6.5-7.8 (Gross and Werner 1978). It is found from sea level to
8,000 feet (2,440 m) elevation.
åÊ
|
|
WHERE DID IT COME FROM AND HOW IS IT SPREAD?
|
Common mullein is native to Asia, but it
probably was introduced to the United States from Europe. It was valued for its
medicinal properties and has been carried with immigrants throughout the world.
It has been used as a remedy for coughs and lung diseases, diarrhea, burns, and
earaches (Mitich 1989). It probably was introduced several times into North
America as a medicinal herb as well as accidentally. The earliest recorded
intentional introduction was in the 1700s in the Blue Ridge Mountians of
Virginia (Gross and Werner 1978). It apparently naturalized and spread rapidly,
for it was erroneously described as a native by Eaton (1818) and was present as
far west as Michigan in 1839 (Gross and Werner 1978). It was first recorded in
California in 1880 as being widely naturalized in old fields in Siskyou County
(Watson 1880). It spreads by prodigious seed production and maintains its
presence by long-lived seeds in the soil. Its seeds have no specialized
structures for long-distance dispersal by wind or animals. Movement of soil for
highway and building construction may have assisted in its dispersal.
åÊ
|
|
WHAT PROBLEMS DOES IT CAUSE?
|
Common mullein is not a weed of agricultural
crops, as it cannot tolerate cultivation. It is, however, thought to serve as a
host for insects that are themselves economic pests, such as the mullein leaf
bug, a pest of apples and pears in the eastern United States and Canada (Maw
1980). Common mullein is not often a significant weed of most wildlands and
natural areas, as it is easily crowded out by grasses or other competing
vegetation. It is a problem, however, in the sparsely vegetated soils of the
eastern Sierra Nevada. In moist meadows and drainages near Mono Lake and Owens
Valley, common mullein can become abundant and has invaded pristine meadows with
undisturbed soils, displacing native herbs and grasses. It has also been
observed to rapidly establish following forest fires in the western Sierra
Nevada. High densites of rosettes appear to prevent the reinvasion of native
herbs and grasses in burned areas, but eventually these give way to a developing
shrub canopy. In this situation, mullein appears to disrupt the normal sequence
of ecological succession.
åÊ
|
|
HOW DOES IT GROW AND REPRODUCE?
|
(click on photos to view larger image)
|
Common mullein reproduces solely by seed. Flowering occurs in summer, usually July through October in the eastern Sierra and June to August in the Sacramento and San Joaquin valleys. Flowers are borne on an inflorescence that occurs as an upward extension of the stalk. Individual flowers are short-lived, opening before dawn and closing
|
|
by mid-afternoon the same day. Flowers are visited by a diverse array of
insects, but only the short- and long-tongued bees are effective pollinators.
Flowers are also self-fertile, with self-fertilization occurring when the flower
closes at the end of the day if cross-pollination has not occurred (Gross and
Werner 1978).
Seeds are contained in a capsule with two
cells. Field studies show that single plants produce 200 to 300 capsules with
500 to 800 seeds per capsule. Thus, seed production can be 100,000 to 240,000
seeds per plant. When dry, the capsule splits open and releases the seeds. Seeds
are not adapted to dispersal by wind or animals and usually fall to the ground.
Field studies report that seeds will disperse as far as eleven meters, but 75
percent fall within one meter of the parent plant (Gross and Werner 1978).
Common mullein seeds do not
appear to undergo dormancy or require a period of after-ripening. They
germinate rapidly under appropriate environmental conditions (Baskin and Baskin
1981). Seed germination can occur in continuous darkness (e.g., when buried) or
in light. High germination rates in darkness are restricted to
relatively hightemperatures (>30 degrees C). In contrast, high germination rates
were observed at 0 degrees C in darkness alternating with 35-40 degrees C in
light (Semenza et al. 1978). This indicates that
germination is possible on soil surfaces where extreme diurnal fluctuations occur. Despite observations of seed germination in darkness in the laboratory, field studies of buried seeds show low germination rates(
Generally, only those seeds
at or near the soil surface will germinate (Semenza et al. 1978). Mullein seeds can
survive and remain viable for thirty-five to 100 years when buried (Gross and Werner
1978, Baskin and Baskin 1981). The presence of mullein plants immediately following
soil disturbance is likely a result of the presence of a seedbank rather than dispersed
seeds. In California common mullein seeds usually germinate in spring following snowmelt
and in fall with the onset of rains.
Common mullein is a usually a biennial, forming a taproot and a
rosette in the first year and a flowering stalk in the second year. Rosettes
consist of a whorl of leaves from the root crown clustered at the soil surface.
In the eastern Sierra Nevada, however, it can grow as a biennial or as a winter
annual. If seed germinates in spring, the plant will remain a rosette through
the first growing season and the following winter. It will then bolt in spring
and flower in summer. If seed germinates in fall, the plant will enter winter as
a rosette and bolt the following spring (Semenza et al. 1978). Regardless of its
flowering pattern, common mullein spends the first half of its life as a
rosette, producing a deep taproot before sending up a three- to six-foot stalk
and producing flowers. Flowers may be produced until the first frost or snowfall
in late fall.
åÊ
|
|
HOW CAN I GET RID OF IT?
|
The best method for controlling common
mullein depends on the size of the infestation, the topography of the site, and
the resources available. Timing is critical for efficient control, and follow-up
is essential.
åÊ
|
|
Physical control:
|
Manual/mechanical methods: Perhaps the most effective method of
controlling common mullein is to cut plants with a weed hoe. Plants will not
resprout if cut through the root crown below the lowest leaves (Gross and Werner
1978). Removing rosettes with a hand hoe can be easily accomplished by workers
trained to recognize the plant. Hand hoeing can be selective and effective, and
two workers may clear up to twenty acres of mullein in a few hours. Bolted
plants can also be removed with a weed hoe. Sometimes bolted plants can be
pulled out of sandy soil, especially following heavy rain. If plants have begun
to set seed, cut off the flowering racemes with pruning shears just below the
lowest seed pods and collect them in a bag to prevent seeds from being released
during the hand removal operation. A second or third weeding may be necessary.
Mowing appears to be ineffective, as plants cut above the root
crown do not die. Rather, the basal rosette will continue to enlarge, then later
bolt and flower. Clipping the terminal flower stalk will not prevent flowering,
but will cause increased growth of axillary branches, which will produce flowers
later (Gross and Werner 1978).
Prescribed burning: Burning kills bolted plants and appears to
kill rosettes, but creates open areas for reinfestation from seed germination.
Individual bolted plants can be killed using a flame thrower, but its use is to
be avoided during fire season.
åÊ
|
|
Biological control:
|
Insects and fungi: No insects or diseases
have been approved for introduction as biological control agents against common
mullein in North America. A curculionid weevil, Gymnaetron tetrum Fab.,
was accidentally introduced into Canada from Europe (Maw 1980). This weevil is
specific to common mullein and is considered one of two natural enemies
significantly impacting the plant in Europe. The larvae feed on seeds and other
tissues in the seed capsules. Larvae are able to destroy all seeds in a capsule
when present; however, usually not all seed capsules are infested. Gross and
Werner (1978) report that up to 50 percent of seeds may be destroyed by the
larval feeding of this weevil. Over time, G. tetrum has spread into
California and has been collected from mullein plants throughout northern and
eastern California since 1942. While its impact has not been investigated in
California, it is unlikely to have much impact on common mullein populations.
Despite the rate of seed destruction, too many seeds remain for it to have much
effect in controlling common mullein populations.
Grazing: Grazing animals generally will not eat mullein because
of its hairy leaves (Whitson 1992).
åÊ
|
|
Chemical control:
|
Common mullein is difficult to control with herbicides because
the thick hairs on the leaves prevent the herbicide from reaching and
penetrating the leaf surface. A surfactant is recommended for all liquid
herbicides used to control this plant.
Zamora (1993) compared the effectiveness of several herbicides
to control common mullein along a roadside in Montana. Herbicides were applied
with a backpack sprayer calibrated to deliver 20 gal/acre at 42 psi. Treatments
were applied to late rosette and bolting plants in late May. Plant height and
number of plants surviving to flower were recorded at the end of July. Of the
three compounds available in California, 2,4-D provided 66 percent kill at 1.9
lbs/acre; height of standing plants averaged nineteen inches. Glyphosate
provided 100 percent kill; height averaged nine inches. Sulfometuron (as Oustå¨)
also provided 100 percent kill; average plant height was six inches. For
comparison, in the unsprayed control areas, all plants survived to flowering
(zero mortality); average plant height was thirty-eight inches (1 m).
Another control method, recently developed by a forest weed
manager, is to spray each rosette with glyphosate by putting the spray nozzle
into the center of the rosette (DiTomaso, pers. comm.). The applicator touches
the plant with the spray nozzle and gives it one good squirt. The key is to
ensure that the herbicide penetrates the region of the plant where the growing
point is located. If the nozzle is off-center, this method does not work. Only
seedlings and rosettes are susceptible using this method. In treating individual
plants, it is recommended that a dye be used in the herbicide mixture to mark
treated plants and prevent re-treatment.
åÊ
|